2024: Volume 3, Issue 2
Article In Press
 Abstract
Abstract  PDF
PDFRepurposing N-acetylcysteine: Unveiling Its Potential for Memory Enhancement in Animal Model
Huma Ikram1,*, Sania Ajaz1, Mahwish Iftikhar1, Shehar Bano1, Sadaf Zaheer1, Atufa Shabbir1, Darakhshan Jabeen Haleem1,2
1Neurochemistry and Biochemical Neuropharmacology Research Unit, Department of Biochemistry, University of Karachi, Karachi-75270, Pakistan
2Neuroscience Research Laboratory, Dr. Panjwani Center for Molecular Medicine and Drug Research, University of Karachi, Karachi-75270, Pakistan
*Corresponding author: Huma Ikram, Department of Biochemistry, University of Karachi, Karachi-75270, Pakistan, Tel: +92-322-2697307, E-mail: [email protected]
Received Date: July 14, 2024
Publication Date: November 01, 2024
Citation: Ikram H, et al. (2024). Repurposing N-acetylcysteine: Unveiling Its Potential for Memory Enhancement in Animal Model. Nutraceutical Res. 3(2):09.
Copyright: Ikram H, et al. © (2024).
ABSTRACT
Objectives: N-acetylcysteine (NAC) is a well-known antioxidant and mucolytic agent with emerging evidence suggesting its neuroprotective properties. This study investigates the potential of NAC to enhance memory in an animal model, exploring its repurposing for cognitive enhancement. Methods: Adult male Albino Wistar rats weighing 200±20g were orally administered NAC at the dose of 25, 50 and 100mg/kg. On day 0, rats underwent training in the Morris Water Maze (MWM) and familiarization in the Novel Object Recognition (NOR) test. Following administration of water or respective NAC doses, memory performance was assessed using the MWM test, measuring acquisition, retention and consolidation. The NOR test was also conducted to evaluate recognition memory. Additional behavioral assessments included the open field, Skinner’s box, light-dark activity box, elevated plus maze, and forced swim test were also performed. Rats were then decapitated; brain samples were isolated and stored at -70°C until analysis by HPLC-EC. Results: NAC administration showed a dose-dependent improvement in memory performance in both the MWM and NOR tests. Behavioral activities indicated enhanced cognitive function and reduced anxiety-like behaviors post-NAC administration. Neurochemical analysis of brain samples revealed changes in dopamine and serotonin metabolism. Conclusion: NAC demonstrates capacity for memory enhancement and cognitive improvement, with effects measured via several behavioral assessments. Findings may help in extending therapeutics in cognitive disorders.
Keywords: N-acetylcysteine, Memory Enhancement, Neuroprotection, Oxidative Stress
INTRODUCTION
N-acetylcysteine (NAC) is usually used as a mucolytic agent in treating chronic obstructive pulmonary disease and cystic fibrosis [1]. It is a popular antidote for paracetamol (acetaminophen) overdose, because it restores intracellular glutathione, an essential antioxidant that neutralizes toxic metabolites of paracetamol, preventing liver damage [2]. NAC is known for its extremely good protection profile, with no toxicity typically related to large doses of other drugs/ substances. The antioxidant properties of N-acetylcysteine (NAC) offer a biochemical foundation in treating diverse situations associated with oxidative stress and inflammation [3]. These properties enable NAC to combat the harmful results of reactive oxygen species (ROS) and reduce inflammatory responses, making it a promising healing agent for neurodegenerative, cardiovascular, and psychiatric disorders [4]. NAC's large therapeutic capability is supported through its capacity to modulate redox potential and inflammatory pathways, emphasizing its role in addressing pathological conditions wherein oxidative stress are potential contributing factors [5].
NAC is a thiol compound recognized for its capability to stimulate the property of cytosolic enzymes critical for the regeneration of glutathione (GSH), a critical antioxidant inside the body. Additionally, NAC acts as a direct precursor for GSH synthesis, thereby improving the body's antioxidant potential [6]. Beyond its well documented antioxidant role, NAC has the precise capability to cross the blood-brain barrier. These functions may allow NAC induced increased GSH levels within the brain, as suggested in preclinical trials, thereby exerting its protecting effects in the central nervous system. Moreover, NAC has been shown to offer a whole lot of neuroprotective effects in numerous neurological preclinical models. It enables safety in reaction to oxidative stress via neutralizing reactive oxygen species and lowering oxidative damage to neurons [7,8]. NAC also has the functionality to chelate heavy metals, that could make contributions to neurotoxicity. By influencing glutamate neurotransmission, NAC permits stability of excitatory and inhibitory neurotransmitters in the mind, which is crucial for regular cognitive functions. Additionally, NAC has been mentioned to decrease inflammatory markers, thereby decreasing neuroinflammation, a common observed in diverse neurological disorders brought about by oxidative stress [9].
NAC's functionality to inhibit apoptosis, or programmed cell death, in addition underscores its neuroprotective ability. It enhances neurogenesis which is important for preserving cognitive abilities and repairing brain damage. Furthermore, NAC promotes neuronal survival through supporting general health and characteristics of neurons. These numerous mechanisms of action spotlight NAC's restoration capacity in treating a vast variety of neurological conditions, as evidenced through preclinical trails [10,11]. Glutathione (GSH) performs a pivotal role in the antioxidant action of NAC because of the fact it is a robust antioxidant and a critical cofactor for several antioxidant enzymes that defend cells from free radical damage. By replenishing GSH levels, NAC enhances cell protection in response to oxidative stress, thereby mitigating damage from reactive oxygen species (ROS) [12]. As previous studies suggested that 2-cyclohexene-1-1 (CHX) can impair short-term spatial memory in rats and mice, however NAC has the capacity to restore GSH levels and alleviate those cognitive deficits [13].
Due to its neuroprotective results, NAC may also offer protection in the direction of a whole lot of neurodegenerative situations consisting of Parkinson's, Alzheimer's, stroke, and multiple sclerosis (MS). NAC's efficacy in these conditions may be attributed to its role as a precursor to glutathione, mediating its anti-inflammatory and antioxidant capabilities. As a precursor to GSH, NAC helps maintain redox stability and supports the function of antioxidant enzymes, that is critical in neuroprotection [14]. Additionally, its anti-inflammatory results help to lessen neuroinflammation, that is a common characteristic of many neurodegenerative illnesses. Animal studies have set up tremendous promise for NAC in restoring cognitive overall performance, even in conditions where the pathophysiology of β-amyloid, a hallmark of Alzheimer's diease, remains unchanged. This shows that NAC's therapeutic potential goes beyond its antioxidant potential, probably related to modulation of various pathways implicated in cognitive feature and neurodegeneration [15]. Furthermore, research shows that NAC, when used as an adjuvant therapy, can also help to increase the lifespan of dementia patients. Its capability to maintain antioxidant defenses and reduce toxicity may additionally slow down the progression of neurodegenerative strategies, thereby enhancing fundamental cognitive profile of these patients [16]. This multifaceted method highlights NAC's capability as a versatile healing agent for the management of neurodegenerative diseases and cognitive decline. NAC can be administered orally and optimum plasma concentration (Cmax) of NAC is received after 1-2 hours after oral administration because of fast absorption in the gastrointestinal system. NAC has an anticipated 6 hour t½ [17]. Understanding the mechanisms via which NAC exerts its outcomes on memory ought to pave the way for its use as a restoration agent in the remedy of cognitive impairments related to neurodegenerative disorders.
Serotonin and dopamine play significant roles in the memory enhancement consequences of NAC. NAC affects the serotonergic levels that would affect stress, anxiety and cognitive abilities, thereby indirectly assisting memory formation [18]. NAC has been reported to increase dopamine release and inhibit the adverse consequences of 6-OHDA on the expression of tyrosine hydroxylase [19]. Dopamine is a critical neurotransmitter concerned in diverse cognitive methods, along with learning, memory, and cognitive features. NAC also protects dopaminergic neurons from oxidative pressure, that's crucial for preserving synaptic plasticity and neurotransmission [20]. By enhancing dopamine signaling, NAC may additionally enhance memory formation and retrieval, thereby contributing to its capability as a therapeutic agent for cognitive enhancement.
The present study was designed to explore the efficacy of NAC in enhancing memory and cognitive functions in an animal model, particularly its effects on dopamine and serotonin metabolism. Using behavioral assessments and neurochemical evaluation, we monitored NAC's potential as a cognitive enhancer. Findings intend to offer implications for the improvement of treatment strategies for cognitive disorders.
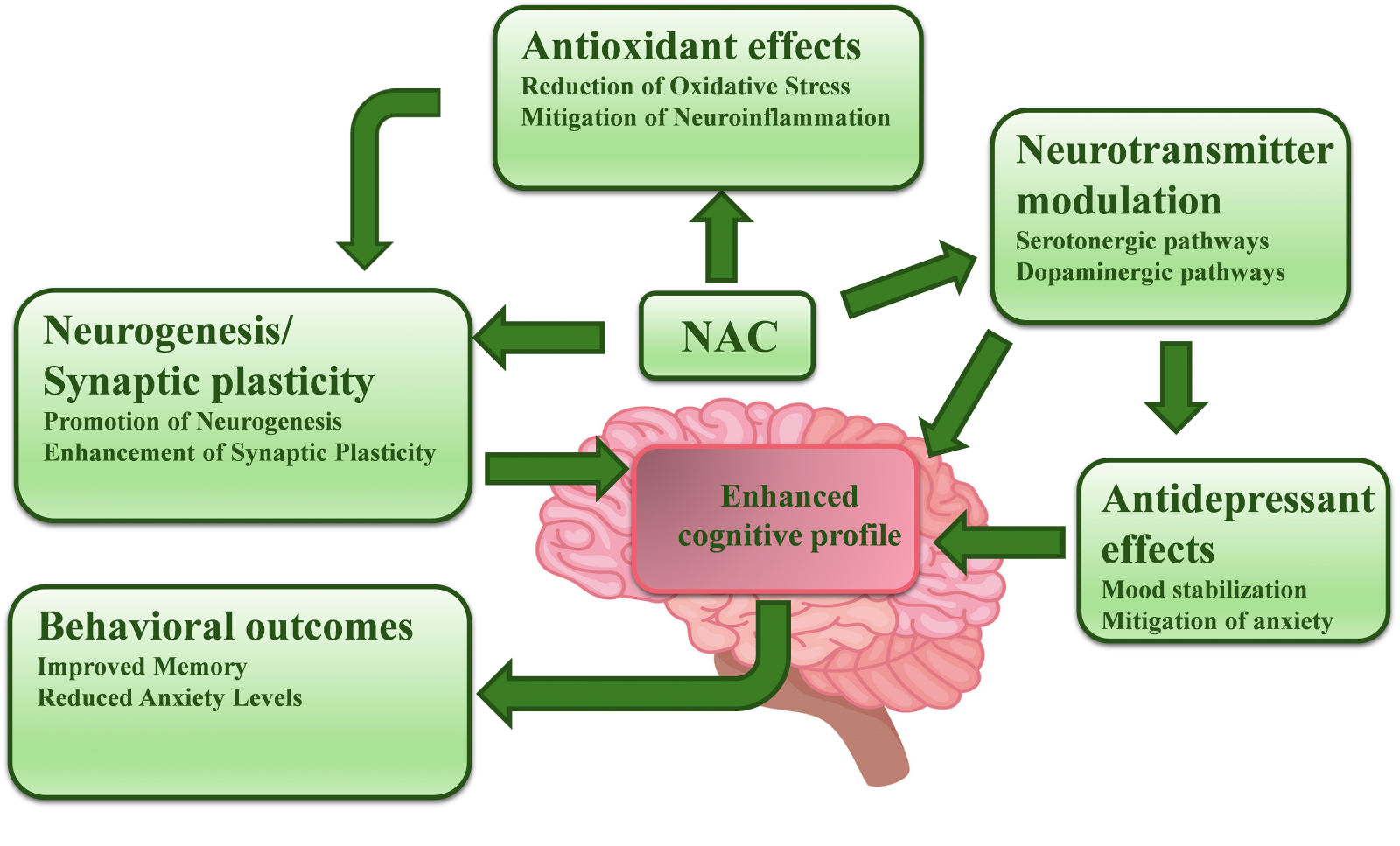
Proposed mechanisms of N-acetylcysteine (NAC) for memory enhancement
MATERIALS AND METHODS
Animals
Albino-Wistar rats (200±20 g) purchased from HEJ Research Institute of Chemistry, University of Karachi, Pakistan were housed individually in the perspex housing cages. Animals were placed in environmentally controlled room under a 12:12 h light/ dark cycle (lights on at 6:00 hr). An acquisition phase of three days was allowed for familiarization. All experiments were performed in accordance with NIH suggestions and permitted by means of the Institutional Ethics and Animal Care Committee (approval number: KU-12072023).
Drug and Doses
N-acetylcysteine (Sigma, St. Louis, USA) was dissolved in saline (0.9% NaCl solution) and administered orally at the dose of 25mg/kg, 50mg/kg and 100mg/kg to respective rats. Drug solution was freshly prepared each day before starting the experiment. Water was administered to the control rats orally at the dose of 1ml/kg.
Experimental Protocol
Twenty-four male Albino Wistar rats were randomly assigned to four groups each containing six rats: (i) water, (ii) acetylcysteine (25mg/kg), (iii) acetylcysteine (50mg/kg) and (iv) acetylcysteine (100mg/kg) treated rats. Doses were selected in line with the other studies reporting no toxicity within this range [21]. On day 0 (training day) rats were trained in Morris water maze with a cut off time of 5min. Rats were familiarized with the familiar object in novel objection recognition test only with a cut off time of 10min. Rats failing to reach the platform within 1 min were introduced to the platform and allowed to explore it for 30 secs and the rats with no recognition of the object were made to sniff the object to get familiarized. On day 1, rats were administered with water (1ml/kg) or acetylcysteine (respective dose). Memory revival was monitored through Morris water maze test. Parameters included; training (24hr pre-injection), acquisition (1hr post-injection), retention (24hr post-injection) probe test/ consolidation (48hr post-injection). and novel objection recognition test was also performed to assess recognition memory. Activities in open field, Skinner’s box, light dark activity box, elevated plus maze and forced swim test were also monitored post acetylcysteine administration. Rats were decapitated 24hr post administration. Brain samples were collected and stored at -70°C until neurochemical analysis by HPLC-EC.
Behavioral Procedures
Activities in the familiar environment of Skinner’s box
Transparent Perspex cages (26×26×26 cm) with a saw dust-covered floor were used to monitor activity in a familiar environment. Rats were placed individually in these cages to become familiar with the environment. Fifteen minutes later, the animals were administered drugs or water. The number of cage crossings was counted 5 min postinjection for 10 min [22].
Activities in a novel environment of open field
A square area (76×76 cm) with walls 42 cm high was used to monitor activity in a novel environment. The floor of the apparatus was divided by lines into 25 squares of equal size. Animals were injected with drug or vehicle and placed in the central square of the open field immediately after the injection. The numbers of squares crossed with all four paws were counted for 5 min [23].
Elevated plus maze test
Procedure was same as described earlier [24]. Elevated plus-maze is a cross shaped maze that has two open arms and two close arms enclosed by sides, but with an open roof. The entire maze is elevated 50cm above the floor. Rodent’s unconditioned aversion to light and open spaces contribute to its effectiveness as a test for anxiety. Test involves placement of animal in the center of maze and observing the number of entries and time spent in open/fear inducing arm.
Light dark activity box
Specifically designed two Perspex compartments of equal dimensions (26x26x26 cm) were used to monitor the activity. One compartment was transparent and other was black walled with an entry between them. Experiment was conducted in a separate room. To determine light and dark field activity, an animal was taken out from home cage and placed for the first time in the light compartment. Number of entries in light compartment and time spent in the light and dark compartments were monitored for 5 minutes [25].
Forced swim test
Specifically designed two Perspex compartments of equal dimensions (26x26x26 cm) were used to monitor the activity. One compartment was transparent and other was black walled with an entry between them. Experiment was conducted in a separate room. To determine light and dark field activity, an animal was taken out from home cage and placed for the first time in the light compartment. Number of entries in light compartment and time spent in the light and dark compartments were monitored for 5 minutes [26].
Morris water maze test
The procedure of the Morris water maze test was essentially same as described before [27]. Animals were trained to locate the hidden platform in the water maze 1hr post injection. After training, animals were placed back in their home cages. Learning acquisition was monitored as latency to locate the platform. Tests for memory acquisition were monitored for 1hr post training and immediately after drug administration. While memory retention and extinction were monitored for 1hr and 6hr post injection respectively. Probe test was performed without platform and time taken to reach the target quadrant was monitored. Memory reconsolidation was monitored 24hr post previous injection and immediately after next drug administration.
Novel object recognition test
For the novel object recognition test the apparatus used was an open arena of (50 × 50) with 42 cm high walls. Procedure was same as described earlier [28]. To saturate it with olfactory stimuli, cleaning of box was not allowed throughout the experiment. The objects to be discriminated were two glasses; one was filled with white cement (used as familiar object) to make it heavy enough so that rats could not be able to move it, and a metallic colored object (used as a novel object). The size of the objects was 2.5 times the size of the rat so that the rat could easily sniff it. During the first training session individual rat was permitted to explore the open field arena for 10min, so that the animal was familiarized to the environment. After a delay of 24 hours second training session was performed. During this session two similar novel objects were placed in testing area and animals were allowed to explore them for 10 min. After the delay of 24 hours the retention test was performed in which the animal was positioned back into the similar environment the only difference is that one of the familiar objects (used in training session was now replaced by a novel object and each animal was given a maximum of 10 min to accumulate 30 seconds of object exploration.
Statistical Analysis
Results are given as means±SD. Analysis of the data was performed by one-way/ two-way ANOVA, wherever applicable. Post hoc comparisons were done by Tukey’s test. Values of p<0.01 were considered statistically significant.
RESULTS
Figure 1 shows dose dependent effects of NAC on exploratory activities in familiar as well as novel environments. Analysis of the data on activities in familiar environment (Figure 1a) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 102.56; p= 0.0001) on exploratory activities. Post hoc analysis by Tukey’s test showed increased (p<0.01) activity in rats administered with NAC at all three dose as compared to 0mg/kg treated rats. Analysis of the data on activities in novel environment (Figure b) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 92.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) activity in rats administered with NAC at the dose of 50mg/kg as compared to both 0mg/kg as well as 25mg/kg NAC treated rats.
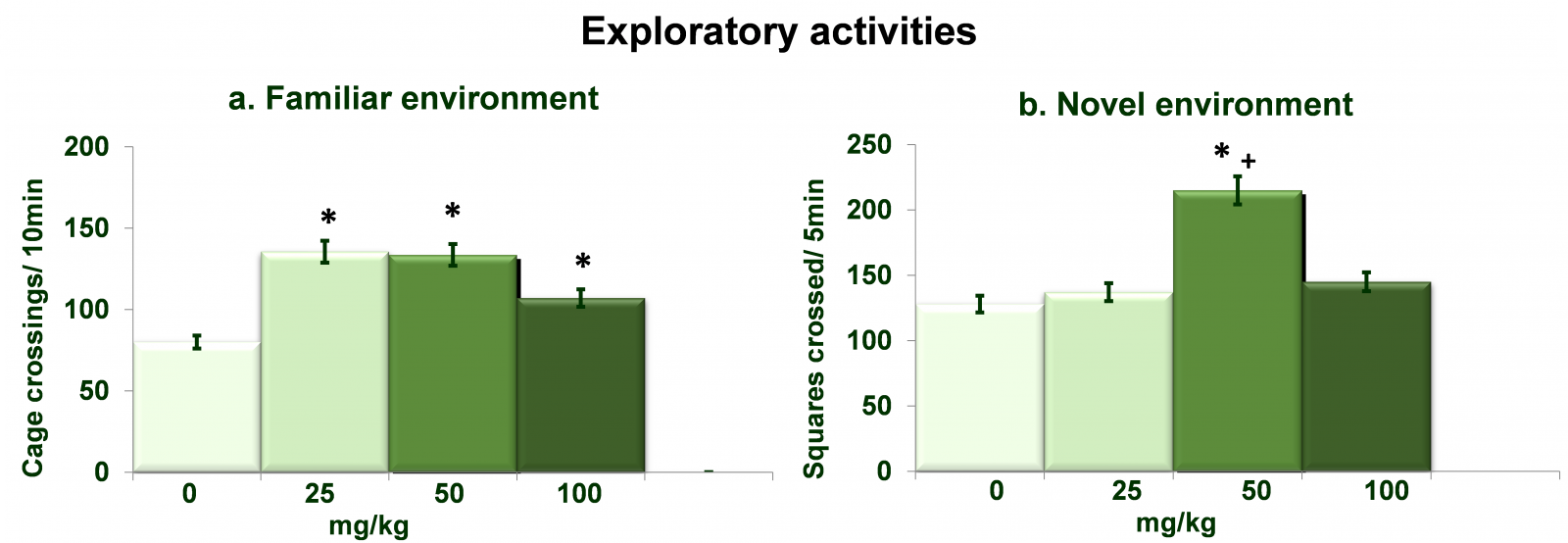
Figure 1. Dose dependent effects of NAC on exploratory activities in familiar as well as novel environments. Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to 0mg/kg NAC treated rats; +p<0.01 as compared to 25mg/kg NAC treated rats, following one-way ANOVA.
Figure 2 shows dose dependent effects of NAC on elevated plus maze activity. Analysis of the data on entries in arms (Figure 2a) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 92.36; p= 0.0001), arms (df= 1,40; F= 68.23; p= 0.0001) and interaction between the two (df= 3,40; F= 92.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) entries (in open arm) at all three doses of NAC as compared to respective entries (in close arm) values as well as compared to 0mg/kg NAC treated rats. While entries (in open arm) were also greater (p<0.01) in 50mg/kg NAC treated rats as compared to 25mg/kg NAC treated rats. Entries (in closed arm) at all three doses of NAC as were decreased (p<0.01) as compared to respective entries (in close arm) by 0mg/kg NAC treated rats. Analysis of the data on time spent in arms (Figure 2b) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 96.32; p= 0.0001), arms (df= 1,40; F= 86.49; p= 0.0001) and interaction between the two (df= 3,40; F= 68.52; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) time spent (in open arm) at all three doses of NAC as compared to respective time spent (in close arm) values as well as compared to 0mg/kg NAC treated rats. While time spent (in open arm) was also greater (p<0.01) in 50mg/kg NAC treated rats as compared to 25mg/kg NAC treated rats. Time spent (in closed arm) at all three doses of NAC as were decreased (p<0.01) as compared to respective time spent (in close arm) by 0mg/kg NAC treated rats.
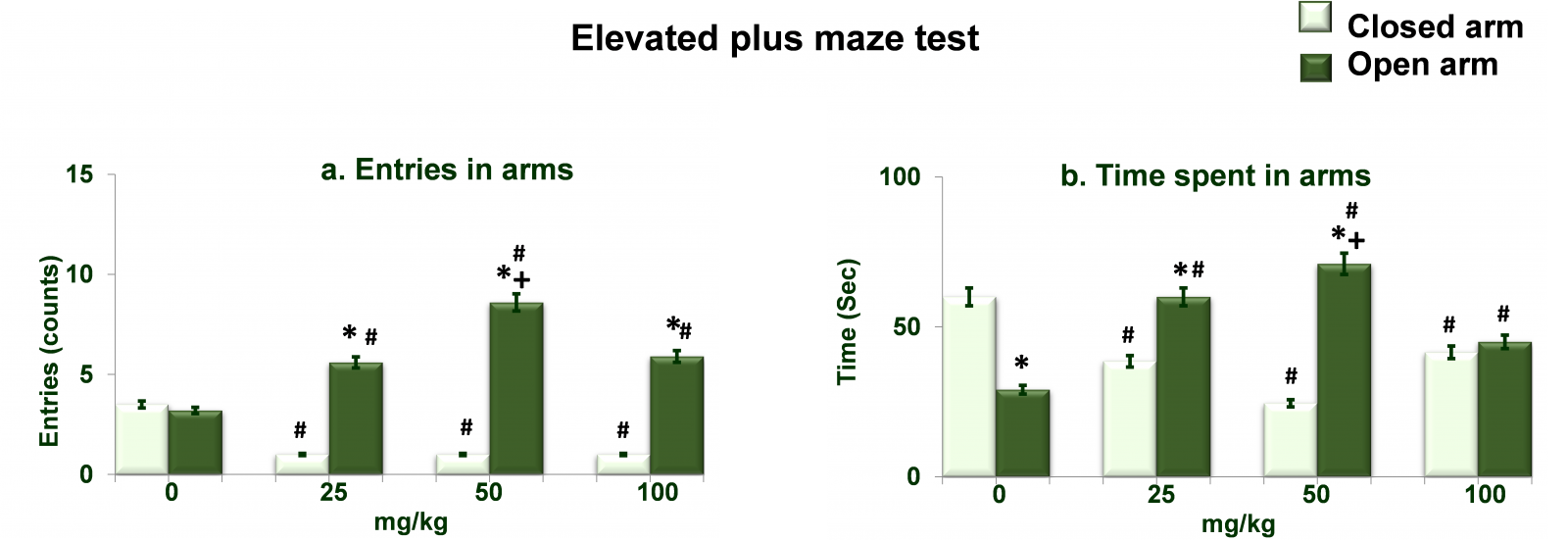
Figure 2. Dose dependent effects of NAC on elevated plus maze test. Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to closed arm; +p<0.01 as compared to 25mg/kg NAC treated rats, #p<0.01 as compared to 0mg/kg NAC treated rats, following two-way ANOVA.
Figure 3 shows dose dependent effects of NAC on light dark box activity. Analysis of the data on entries in compartments (Figure 3a) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 68.36; p= 0.0001), arms (df= 1,40; F= 82.16; p= 0.0001) and interaction between the two (df= 3,40; F= 102.56; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) entries (in light compartment) at all three doses of NAC as compared to respective entries (in dark compartment) values as well as compared to 0mg/kg NAC treated rats. While entries (in light compartment) in 50mg/kg NAC treated rats were greater (p<0.01) than 25mg/kg NAC treated rats. Analysis of the data on time spent in compartments (Figure 3b) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 56.25; p= 0.0001), arms (df= 1,40; F= 116.23; p= 0.0001) and interaction between the two (df= 3,40; F= 84.23; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) time spent in light compartment in 0mg/kg treated rats as compared to time spent in dark compartment by same rats. Time spent in light compartment was increased (p<0.01) at 25mg/kg and 50mg/kg NAC treated rats as compared to time spent in dark compartment by same rats as well as in comparison to 0mg/kg treated rats. Time spent in light compartment in 50mg/kg treated was also more (p<0.01) 25mg/kg NAC treated rats. Time spent in light compartment was increased (p<0.01) at 100mg/kg NAC treated rats as compared to 0mg/kg treated rats.
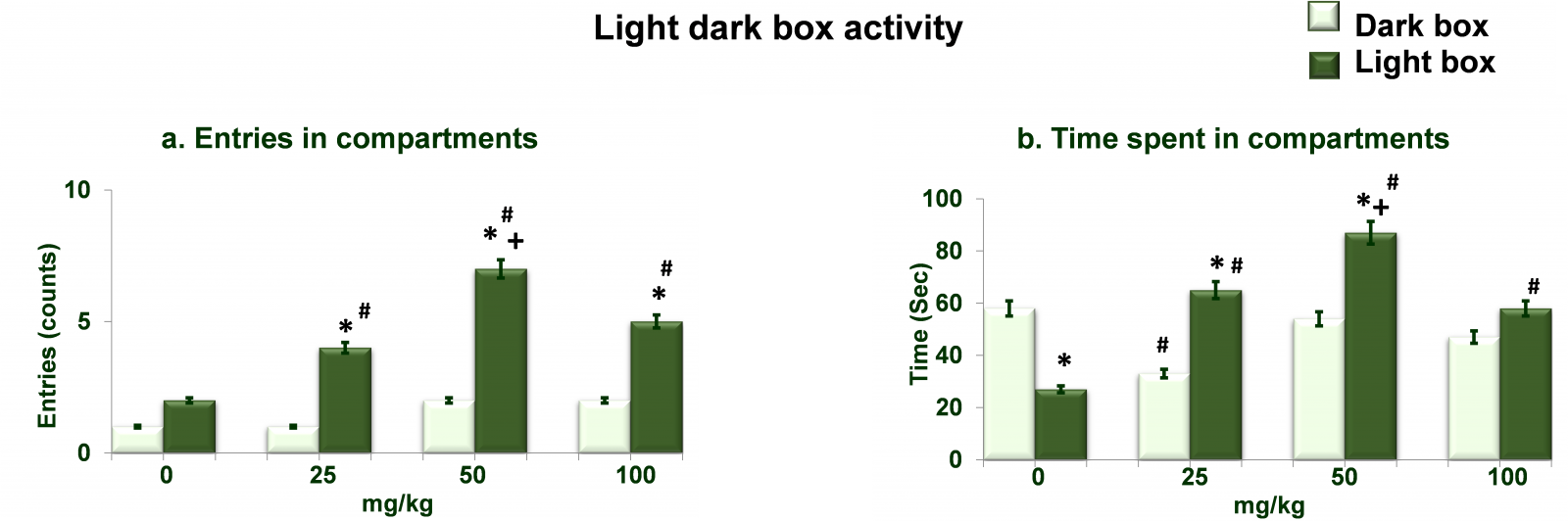
Figure 3. Dose dependent effects of NAC on light dark box activity. Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to dark compartment; +p<0.01 as compared to 25mg/kg NAC treated rats, #p<0.01 as compared to 0mg/kg NAC treated rats, following two-way ANOVA.
Figure 4 shows dose dependent effects of NAC on forced swim test. Analysis of the data on number of jumps (Figure 4a) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 75.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) jumps in rats treated with all three doses of NAC as compared to 0mg/kg NAC treated rats. In 50mg/kg and 100mg/kg NAC treated rats number of jumps were also increased (p<0.01) as compared to 25mg/kg treated rats. Analysis of the data on struggle time (Figure 4b) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 94.82; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) struggle time in rats treated with 50mg/kg doses of NAC as compared to 0mg/kg as well as 25mg/kg NAC treated rats. Data on flat body posture (Figure 4c) as analyzed by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 91.25 p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) flat body posture in rats treated with 50mg/kg doses of NAC as compared to 0mg/kg as well as 25mg/kg NAC treated rats.
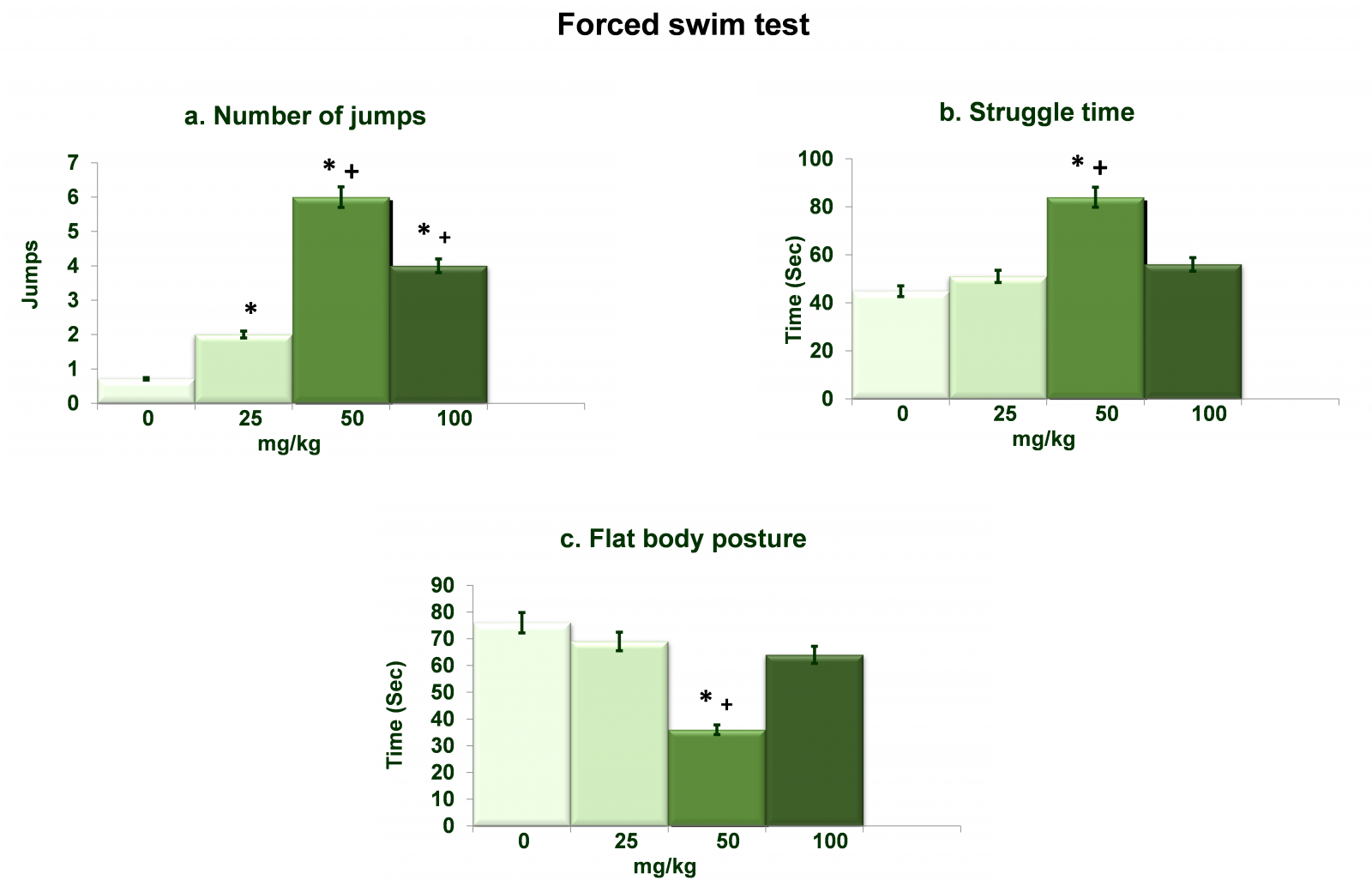
Figure 4. Dose dependent effects of NAC on forced swim test. Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to 0mg/kg NAC treated rats; +p<0.01 as compared to 25mg/kg NAC treated rats, following one-way ANOVA.
Figure 5 shows dose dependent effects of NAC on Morris water maze test (time taken to reach platform). Analysis of the data on memory acquisition (Figure 5a) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 96.32; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) time taken to reach platform in rats treated with 50mg/kg NAC as compared to respective 0mg/kg as well as 25mg/kg NAC treated rats. Analysis of the data on memory consolidation (Figure 5b) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 52.36; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) time taken to reach platform in rats treated with 50mg/kg NAC as compared to respective 0mg/kg as well as 25mg/kg treated rats. Analysis of the data on memory retention (Figure 5c) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 85.36; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) time taken to reach platform in rats treated with 50mg/kg NAC as compared to respective 0mg/kg as well as 25mg/kg NAC treated rats. Time taken to reach platform was also decreased in rats treated with 100mg/kg NAC as compared to respective 0mg/kg NAC treated rats. Data of the probe test (Figure 5d) as analyzed by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 91.36; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) time taken to reach platform in rats treated with 50mg/kg NAC as compared to respective 0mg/kg as well as 25mg/kg NAC treated rats. Time taken to reach platform was also decreased in rats treated with 100mg/kg NAC as compared to respective 0mg/kg NAC treated rats.
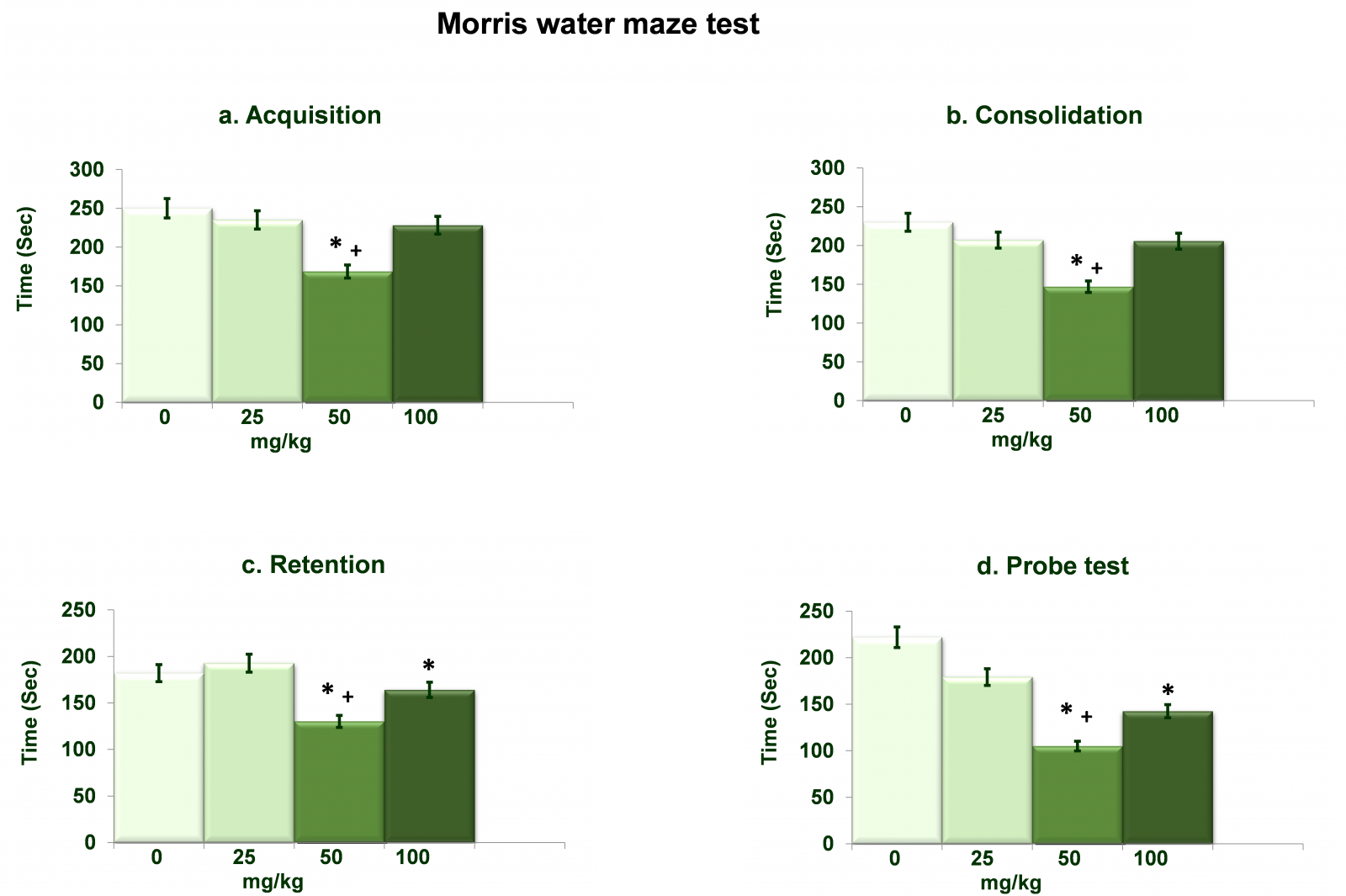
Figure 5. Dose dependent effects of NAC on Morris water maze test (time taken to reach platform). Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to 0mg/kg NAC treated rats; +p<0.01 as compared to 25mg/kg NAC treated rats, following one-way ANOVA.
Figure 6 shows dose dependent effects of NAC on novel object recognition test. Analysis of the data on latency to reach objects (Figure 6a) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 85.36; p= 0.0001), arms (df= 1,40; F= 52.32; p= 0.0001) and interaction between the two (df= 3,40; F= 95.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) time taken for novel object recognition in 0mg/kg as well as 100mg/kg NAC treated rats as compared to time taken for familiar object by the same rats. In 50mg/kg and 25mg/kg treated rats time taken to reach novel object in 25mg/kg as well as 50mg/kg NAC treated rats was decreased (p<0.01) as compared to respective time taken for familiar object as well as 0mg/kg NAC treated rats. Time taken to reach familiar object in 25mg/kg as well as 50mg/kg NAC treated rats was increased (p<0.01) as compared to respective 0mg/kg NAC treated rats. Analysis of the data on number of attempts (Figure 6b) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 79.35; p= 0.0001), objects (df= 1,40; F= 96.36; p= 0.0001) and interaction between the two (df= 3,40; F= 102.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) attempts for novel object at all three doses of NAC treated rats as compared to respective attempts for familiar object. In 50mg/kg and 25mg/kg treated rats attempts for were increased (p<0.01) as compared to respective 0mg/kg NAC treated rats. Attempts for familiar object in 50mg/kg NAC treated rats were increased (p<0.01) as compared to respective 25mg/kg NAC treated rats. Analysis of the data on exploration time (Figure 6c) by two-way ANOVA showed significant effects of different doses of NAC (df= 3,40; F= 105.36; p= 0.0001), objects (df= 1,40; F= 96.32; p= 0.0001) and interaction between the two (df= 3,40; F= 84.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) exploration time for novel object at all three doses of NAC as compared to respective familiar object as well as 0mg/kg NAC treated rats. In 50mg/kg, treated rats exploration time for novel object was greater (p<0.01) than 25mg/kg NAC treated rats as well.
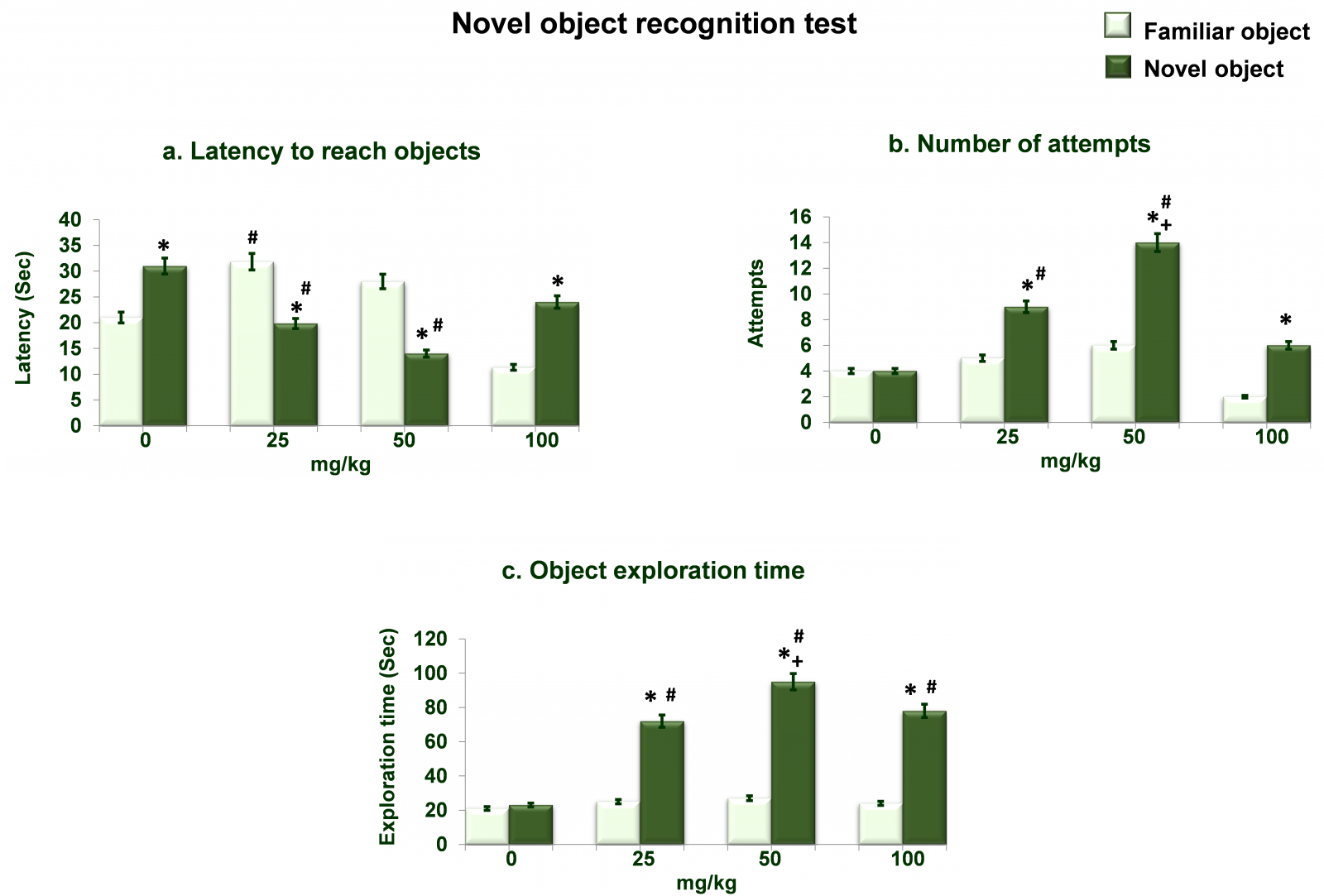
Figure 6. Dose dependent effects of NAC on novel object recognition test. Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to familiar object; +p<0.01 as compared to 25mg/kg NAC treated rats; #p<0.01 as compared to 0mg/kg NAC treated rats following two-way ANOVA.
Figure 7 shows dose dependent effects of NAC on dopamine, 5HT and metabolites in rat whole brain. Analysis of the data on dopamine levels (Figure 7a) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 215.36; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) levels of dopamine at all three doses of NAC as compared to 0mg/kg NAC treated rats. Analysis of the data on DOPAC levels (Figure 7b) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 156.36; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) levels of DOPAC in rats administered with 25mg/kg and 100mg/kg NAC as compared to respective 0mg/kg NAC treated rats. In 50mg/kg treated rats, levels of DOPAC were increased (p<0.01) as compared to 25mg/kg NAC treated rats. Analysis of the data on HVA levels (Figure 7c) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 154.36; p= 0.0001). Post hoc analysis by Tukey’s test showed decreased (p<0.01) levels of HVA at all three doses of NAC as compared to 0mg/kg NAC treated rats. In 50mg/kg and 100mg/kg treated rats, levels of HVA were increased (p<0.01) as compared to 25mg/kg NAC treated rats as well. Analysis of the data on 5HT levels (Figure 7d) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 256.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) levels of 5HT in 50mg/kg NAC treated rats as compared to 0mg/kg and 25mg/kg NAC treated rats. Analysis of the data on 5HIAA levels (Figure 7e) by one-way ANOVA showed significant effects of different doses of NAC (df= 3,20; F= 258.36; p= 0.0001). Post hoc analysis by Tukey’s test showed increased (p<0.01) levels of 5HIAA in 25mg/kg and 50mg/kg NAC treated rats as compared to respective 0mg/kg NAC treated rats.
.png)
Figure 7. Dose dependent effects of NAC on dopamine, 5HT and metabolites (DOPAC: 3,4-Dihydroxyphenylacetic acid; HVA: Homovanillic acid; 5HIAA: 5-Hydroxy indoleacetic acid) in rat whole brain. Values are means±SD (n=6). Significant differences by Tukey’s test: *p<0.01 as compared to 0mg/kg NAC treated rats; +p<0.01 as compared to 25mg/kg NAC treated rats, following one-way ANOVA.
DISCUSSION
The results show that effects of NAC on exploratory activities were dose-dependent, as seen in both the Skinner’s box and open field (Figure 1). At 25mg/kg, NAC-treated rats exhibited a moderate increase in exploratory activity, improving overall cognitive performance and reducing anxiety without causing hyperactivity. At 50mg/kg, exploratory activities were considerably better, suggesting greatest stability among cognitive enhancement and stress reduction, which could be due to effective neurotransmitter modulation. However, at 100mg/kg, even as exploratory activities improved, signs of hyperactivity were observed indicating this dose may also exceed the suggested therapeutic limit for cognitive enhancement. Other studies have reported that NAC improves long-term locomotor characteristics by way of ameliorating oxidative stress-caused neuronal loss, demyelination, leukocyte infiltration and inflammatory mediator expression [29].
The effects of NAC on anxiety-related behaviors were dose-dependent, as shown in the elevated plus maze test and light-dark activity box (Figure 2 & 3). At 25mg/kg, NAC-treated rats showed a moderate reduction in anxiety, spending more time and making more entries in the open arms of the EPM. At 50mg/kg, the anxiolytic effects were significantly enhanced, with rats spending much more time in the open arms and showing increased exploratory behavior. However, at 100mg/kg, although time in the open arms remained high, signs of hyperactivity were observed, suggesting overstimulation. Similar patterns were seen in the light-dark box test, where 25mg/kg resulted in mild anxiety reduction, 50mg/kg showed substantial anxiolytic effects and increased exploration, but 100mg/kg dose led to hyperactivity. These findings highlight that while NAC has anxiolytic effects, particularly at 50mg/kg and higher doses of NAC may lead to overstimulation. This emphasizes the need for dose optimization in therapeutic applications. These anxiolytic outcomes can be secondary to antioxidant results, as different studies have suggested a reduction in chronic oxidative strain through the cooperative consequences of NAC, vitamin D3, and glutathione, thereby achieving a couple of health benefits [30]. NAC has additionally been stated to be well tolerated in type 2 diabetes patients, improving the scientific signs and symptoms of painful diabetic neuropathy because of its antioxidant effects when administered for eight weeks [31].
The effects of N-acetylcysteine (NAC) on depressive-like behavior in the Forced Swim Test (FST) were dose-dependent, affecting immobility time differently at various doses (Figure 4). At 25mg/kg, NAC-treated rats showed a modest reduction in immobility time, indicating a slight antidepressant-like effects likely due to NAC's antioxidant properties and neurotransmitter modulation. At 50mg/kg, the reduction in immobility time was more pronounced, with rats exhibiting significantly more active coping behaviors, such as increased swimming and climbing, suggesting a stronger antidepressant-like effect. This improvement is attributed to enhanced dopaminergic and serotonergic activity. At 100mg/kg, although immobility time was still reduced, signs of hyperactivity, such as excessive swimming and frantic movements, were observed, indicating potential overstimulation at high doses. These findings highlight NAC's antidepressant-like effects, particularly at 50mg/kg, and the importance of dose optimization. It has been reported that NAC as an antioxidant supplement has a potential antidepressant mechanism as assessed in forced swim test and compared with selective serotonin reuptake inhibitors (SSRIs) as standard antidepressant drug. Diminished synaptophysin immunoreactivity in the prefrontal cortex samples of NAC treated rats was like that seen in fluoxetine. Therefore, they reported a similarity in antidepressant profile of NAC and SSRI [32]. Elucidating the role of serotonin in antidepressant mechanisms of NAC is crucial for optimizing treatment strategies and enhancing therapeutic efficacy, particularly in targeting mood regulation pathways. Understanding this relationship can also guide the development of personalized and more effective treatments for depression and related disorders [33-35].
The effects of NAC on cognitive performance in the MWM and NOR tests (Figure 5 & 6) showed a dose-dependent enhancement in memory and learning. In the MWM test, rats treated with 25mg/kg of NAC demonstrated moderate improvements in learning and retention, while the 50mg/kg dose resulted in significantly faster learning and better spatial memory. However, the 100mg/kg dose, although initially beneficial, showed reduced performance efficiency, possibly due to overstimulation. Similarly, in the NOR test, 25mg/kg of NAC improved recognition memory, with the 50mg/kg dose yielding even greater enhancements. The 100mg/kg dose improved recognition memory compared to controls but was less effective than the 50mg/kg dose. The decreased effectiveness at this higher dose could be attributed to potential side effects such as increased hyperactivity, which might distract from focused cognitive tasks. NAC has been reported to improve attention, processing speed, memory, and executive functioning as an adjuvant therapy with guanfacine, in patients with traumatic brain injury [36]. Others have also reported broad spectrum of short-term and long-term cognitive and behavioral improvements in traumatic brain injury patients as evident by resting state MRI [37,38]. It has also been reported that NAC could act as a modulating agent of inflammation in the brain, thereby improving memory [39]. Serotonin plays a crucial role in learning and memory by modulating synaptic plasticity and facilitating the formation and retrieval of memories. Its regulation of mood and anxiety also indirectly supports cognitive processes [40-43]. However, much literature is not available to support direct involvement of serotonin or its receptors in NAC mediated memory enhancement. Elucidating the role of neurotransmitters, particularly serotonin in NAC's memory-enhancing mechanism is essential for optimizing therapeutic strategies and understanding its comprehensive effects on cognitive function. This insight can improve targeted treatments for memory-related disorders.
The results from the present study on the effects of NAC on dopamine and serotonin metabolism in rat brains revealed dose-dependent changes in neurotransmitter levels and their metabolites (Figure 7). At 25mg/kg, NAC moderately increased dopamine and its metabolites (DOPAC and HVA) and slightly elevated serotonin (5HT) and its metabolite (5HIAA), indicating enhanced neurotransmitter activity that supports cognitive functions and mood stability. At 50mg/kg, these effects were more pronounced, with significant increases in both dopamine and serotonin levels, correlating with improved cognitive performance observed in behavioral tests. However, at 100mg/kg, the increases in neurotransmitter levels were not as pronounced as those at 50mg/kg, suggesting possible dysregulation due to overstimulation. This underscores the importance of dose optimization, with 50mg/kg identified as the most effective dose for enhancing dopaminergic and serotonergic activity, thereby supporting cognitive function and mood regulation. Other studies have reported that NAC has a neuroprotective role in SH-SY5Y cells exposed to 6-OHDA by maintaining cell proliferation and decreasing apoptosis. Additionally, it was also validated that NAC treatment increases dopamine release and protects SH-SY5Y cells against 6-OHDA dysregulations on the proteins TH, VMAT2, and α-synuclein [44]. However, it was not related to improvement in memory/ cognitive profile. Apart from memory disorders, NAC has been reported to reveal promising perspectives in preserving the dopaminergic system capability and modulating Parkinson’s and related disorders mechanisms. Although NAC has been shown to act as an antioxidant and (neuro)protector of the brain, it has yet to be acknowledged how this repurposed drug can improve motor symptomatology. Results from a study revealed that NAC could potentially enhanced viability of dopaminergic neurons and may possibly restore dopamine transporter levels [45].
Serotonin (5HT) plays a crucial role in learning and memory by modulating synaptic plasticity, which is essential for the formation and strengthening of neural connections. 5HT influences the activity of key brain regions involved in memory processing, such as the hippocampus and prefrontal cortex [46-48]. It enhances long-term potentiation (LTP), a cellular mechanism underlying learning and memory, thereby facilitating the encoding and retrieval of information [49]. Additionally, serotonin's regulation of mood and emotional states helps cognitive functions by developing a greater conducive environment for memory approaches. Its involvement in neurogenesis, particularly in the hippocampus, similarly underscores its importance in preserving cognitive improvement [50]. In line of the present findings, NAC at a dose of 50mg/kg produced a slight elevation in 5HT and its metabolite 5HIAA, indicating improved serotonergic transmission. This development correlated with the memory-enhancing outcomes of NAC at that dose, suggesting that the enhancement of serotonergic pastime can be a key mechanism underlying the cognitive benefits observed with this treatment. Thus, NAC's capability to boost serotonin levels could appreciably make contributions to its effectiveness in improving learning and memory.
CONCLUSION
This study demonstrated that NAC has prevalent potential for boosting memory and cognitive features in animal models. The administration of NAC at doses of 25mg/kg, 50mg/kg, and 100mg/kg confirmed dose-dependent improvements in numerous behavioral exams, along with the Morris water maze and novel object recognition test. The 50mg/kg dose produced the maximum reported advantages in spatial learning, recognition memory, and stress reduction, indicating the most excellent balance between efficacy and outcomes. Neurochemical analyses through HPLC-EC revealed that NAC administration increased the levels of dopamine and serotonin, as well as their respective metabolites, DOPAC, HVA, and 5HIAA. These findings endorse that NAC's memory-improving effects are likely mediated through better dopaminergic and serotonergic transmission. The elevation of neurotransmitters at the dose of 50mg/kg dose correlated nicely with behavioral improvements, highlighting the role of those pathways in cognitive enhancement. Overall, this study supports the repurposing of NAC as a potential therapeutic agent for memory enhancement. Its capability to modulate key neurotransmitter functions and enhance cognitive features underscores its promise for treating neurodegenerative illnesses and cognitive impairments. The findings from the present study are limited due to the use of an animal model, which may not fully replicate human memory and cognitive processes, potentially affecting the translatability of results. Additionally, future studies focusing the long-term effects of NAC are needed of understanding the sustained impact and safety of prolonged N-acetylcysteine administration.
REFERENCES
- Mokra D, Mokry J, Barosova R, Hanusrichterova J. (2023). Advances in the Use of N-Acetylcysteine in Chronic Respiratory Diseases. Antioxidants (Basel). 12(9):1713.
- Prescott LF. (2024). Paracetamol (acetaminophen) poisoning: The early years. Br J Clin Pharmacol. 90(1):127-134.
- Tieu S, Charchoglyan A, Paulsen L, Wagter-Lesperance LC, Shandilya UK, Bridle BW, et al. (2023). N-Acetylcysteine and Its Immunomodulatory Properties in Humans and Domesticated Animals. Antioxidants (Basel). 12(10):1867.
- Tenório MCDS, Graciliano NG, Moura FA, Oliveira ACM, Goulart MOF. (2021). N-Acetylcysteine (NAC): Impacts on Human Health. Antioxidants (Basel). 10(6):967.
- Jomova K, Raptova R, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, et al. (2023). Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol. 97(10):2499-2574.
- Di Giacomo C, Malfa GA, Tomasello B, Bianchi S, Acquaviva R. (2023) Natural Compounds and Glutathione: Beyond Mere Antioxidants. Antioxidants (Basel). 12(7):1445.
- Sahasrabudhe SA, Terluk MR, Kartha RV. (2023). N-acetylcysteine Pharmacology and Applications in Rare Diseases-Repurposing an Old Antioxidant. Antioxidants (Basel). 12(7):1316.
- Khovarnagh N, Seyedalipour B. (2021). Antioxidant, histopathological and biochemical outcomes of short-term exposure to acetamiprid in liver and brain of rat: The protective role of N-acetylcysteine and S-methylcysteine. Saudi Pharm J. 29(3):280-289.
- Raghu G, Berk M, Campochiaro PA, Jaeschke H, Marenzi G, Richeldi L, et al. (2021). The Multifaceted Therapeutic Role of N-Acetylcysteine (NAC) in Disorders Characterized by Oxidative Stress. Curr Neuropharmacol. 19(8):1202-1224.
- Guo W, Liu K, Wang Y, Ge X, Ma Y, Qin J, et al. (2024). Neurotrophins and neural stem cells in posttraumatic brain injury repair. Animal Model Exp Med. 7(1):12-23.
- Wu W, Liu BH, Xie CL, Xia XD, Zhang YM. (2018). Neuroprotective effects of N-acetyl cysteine on primary hippocampus neurons against hydrogen peroxide-induced injury are mediated via inhibition of mitogen-activated protein kinases signal transduction and antioxidative action. Mol Med Rep. 17(5):6647-6654.
- Zhou J, Terluk MR, Basso L, Mishra UR, Orchard PJ, Cloyd JC, et al. (2020). N-acetylcysteine Provides Cytoprotection in Murine Oligodendrocytes through Heme Oxygenase-1 Activity. Biomedicines. 8(8):240.
- More J, Galusso N, Veloso P, Montecinos L, Finkelstein JP, Sanchez G, et al. (2018). N-Acetylcysteine Prevents the Spatial Memory Deficits and the Redox-Dependent RyR2 Decrease Displayed by an Alzheimer's Disease Rat Model. Front Aging Neurosci. 10:399.
- Adamu A, Li S, Gao F, Xue G. (2024). The role of neuroinflammation in neurodegenerative diseases: current understanding and future therapeutic targets. Front Aging Neurosci. 16:1347987.
- Shahidi S, Zargooshnia S, Asl SS, Komaki A, Sarihi A. (2017). Influence of N-acetyl cysteine on beta-amyloid-induced Alzheimer's disease in a rat model: A behavioral and electrophysiological study. Brain Res Bull. 131:142-149.
- Schwalfenberg GK. (2021). N-Acetylcysteine: A Review of Clinical Usefulness (an Old Drug with New Tricks). J Nutr Metab. 2021:9949453.
- Papi A, Di Stefano AFD, Radicioni M. (2021). Pharmacokinetics and Safety of Single and Multiple Doses of Oral N-Acetylcysteine in Healthy Chinese and Caucasian Volunteers: An Open-Label, Phase I Clinical Study. Adv Ther. 38(1):468-478.
- Chakraborty S, Tripathi SJ, Srikumar BN, Raju TR, Shankaranarayana Rao BS. (2020). N-acetyl cysteine ameliorates depression-induced cognitive deficits by restoring the volumes of hippocampal subfields and associated neurochemical changes. Neurochem Int. 132:104605.
- El-Habta R, Af Bjerkén S, Virel A. (2024). N-acetylcysteine increases dopamine release and prevents the deleterious effects of 6-OHDA on the expression of VMAT2, α-synuclein, and tyrosine hydroxylase. Neurol Res. 46(5):406-415.
- Speranza L, di Porzio U, Viggiano D, de Donato A, Volpicelli F. (2021). Dopamine: The Neuromodulator of Long-Term Synaptic Plasticity, Reward and Movement Control. Cells. 10(4):735.
- Praharaj DL, Anand AC, Acharya SK. (2022). Dosage of N-Acetyl Cysteine in Acute Liver Failure Not Related to Acetaminophen. J Clin Exp Hepatol. 12(2):726-728.
- Ikram H, S Tasneem, S Perveen, R Zakir, DJ Haleem. (2021). Neurochemical and behavioral effects of fluoxetine on midazolam induce dependence in an animal model of addiction. Pak J Pharm Sci. 4(5):1749-1757.
- Ikram H, Haleem DJ. (2021). Attenuation of Apomorphine-Induced Sensitization by Buspirone. Pharmacol Biochem Behav. 99(3):444-450.
- Ikram H, Tasneem S, Perveen S, Haleem DJ. (2020). Neurochemical and Behavioral Effects of Midazolam: A Dose Related Study. Pak J Pharm Sci. 33(1):85-93.
- Ikram H, Atique I, Perveen S, Zakir R, DJ Haleem DJ. (2021). Neurochemical and behavioral effects of lorazepam: A dose related study. Pak J Pharm Sci. 34(1): 85-93.
- Ikram H, Masood R, Syed S, Haleem DJ. (2023). Neuropharmacological studies on repurposed utilization of pioglitazone in learning and memory: A dose related study. Pak J Pharm Sci. 36(4):1159-1167.
- Ikram H, Zakir R, Haleem DJ. (2024). Memory Enhancing and Neuroprotective Effects of Apomorphine in a Rat Model of Dementia. Met Brain Dis. DOI: 10.1007/s11011-024-01372-1.
- Ikram H, Haleem DJ. (2019). Repeated treatment with a low dose of reserpine as a progressive model of Parkinson's dementia. Pak J Pharm Sci. 32(2):555-562.
- Guo J, Li Y, Chen Z, He Z, Zhang B, Li Y, et al. (2015). N-acetylcysteine treatment following spinal cord trauma reduces neural tissue damage and improves locomotor function in mice. Mol Med Rep. 12(1):37-44.
- Mulè S, Ferrari S, Rosso G, Brovero A, Botta M, Congiusta A, et al. (2024). The Combined Antioxidant Effects of N-Acetylcysteine, Vitamin D3, and Glutathione from the Intestinal-Neuronal In Vitro Model. Foods. 13(5):774.
- Sajedi F, Abdi A, Mehrpooya M, Faramarzi V, Mohammadi Y, Sheida F. (2024). Comparison of therapeutic effects of N-Acetylcysteine with pregabalin in improving the clinical symptoms of painful diabetic neuropathy: a randomized, double-blind clinical trial. Clin Diabetes Endocrinol. 10(1):15.
- Memudu AE. (2022). The Efficacy of N-Acetyl-Cysteine (NAC) Supplementation in FST Model for Screening Antidepressants. Basic Clin Neurosci. 13(6):839-854.
- Russell SE, Skvarc DR, Mohebbi M, Camfield D, Byrne LK, Turner A, et al. (2023). The Impact of N-acetylcysteine on Major Depression: Qualitative Observation and Mixed Methods Analysis of Participant Change during a 12-week Randomised Controlled Trial. Clin Psychopharmacol Neurosci. 21(2):320-331.
- Yang C, Bosker FJ, Li J, Schoevers RA. (2018). N-acetylcysteine as add-on to antidepressant medication in therapy refractory major depressive disorder patients with increased inflammatory activity: study protocol of a double-blind randomized placebo-controlled trial. BMC psychiatry. 18(1):279.
- Carollo M, Carollo N, Montan G. (2024). The promise of N-acetylcysteine in the treatment of obsessive-compulsive disorder. CNS Neurosci Ther. 30(2):e14653.
- Khasnavis S, Belliveau T, Arnsten A, Fesharaki-Zadeh A. Combined Use of Guanfacine and N-Acetylcysteine for the Treatment of Cognitive Deficits After Traumatic Brain Injury. Neurotrauma Rep. 5(1):226-231.
- Vedaei F, Newberg AB, Alizadeh M, Zabrecky G, Navarreto E, Hriso C, et al. (2024). Treatment effects of N-acetyl cysteine on resting-state functional MRI and cognitive performance in patients with chronic mild traumatic brain injury: a longitudinal study. Front Neurol. 15:1282198.
- Clark RSB, Empey PE, Kochanek PM, Bell MJ. (2023). N-Acetylcysteine and Probenecid Adjuvant Therapy for Traumatic Brain Injury. Neurotherapeutics. 20(6):1529-1537.
- Vedaei F, Mashhadi N, Zabrecky G, Monti D, Navarreto E, Hriso C, et al. (2023). N-Acetylcysteine Suppresses Microglial Inflammation and Induces Mortality Dose-Dependently via Tumor Necrosis Factor-α Signaling. Int J Mol Sci. 24(4):3798.
- Tortora F, Hadipour AL, Battaglia S, Falzone A, Avenanti A, Vicario CM. (2023). The Role of Serotonin in Fear Learning and Memory: A Systematic Review of Human Studies. Brain sciences, 13(8):1197.
- Bendis PC, Zimmerman S, Onisiforou A, Zanos P, Georgiou P. (2024). The impact of estradiol on serotonin, glutamate, and dopamine systems. Front Neurosci. 18:1348551.
- Farias CP, Leite AKO, Schmidt BE, de Carvalho Myskiw J, Wyse ATS. (2024). The 5-HT2A, 5-HT5A, and 5-HT6 serotonergic receptors in the medial prefrontal cortex behave differently in extinction learning: Does social support play a role? Behav Brain Res. 463:114922.
- Neganova M, Liu J, Aleksandrova Y, Vasilieva N, Semakov A, Yandulova E, et al. (2024). Development of Neuroprotective Agents for the Treatment of Alzheimer's Disease using Conjugates of Serotonin with Sesquiterpene Lactones. Curr Med Chem. 31(5):529-551.
- El-Habta R, Af Bjerkén S, Virel A. (2024). N-acetylcysteine increases dopamine release and prevents the deleterious effects of 6-OHDA on the expression of VMAT2, α-synuclein, and tyrosine hydroxylase. Neurol Res. 46(5):406-415.
- Caridade-Silva R, Araújo B, Martins-Macedo J, Teixeira FG. (2023). N-Acetylcysteine Treatment May Compensate Motor Impairments through Dopaminergic Transmission Modulation in a Striatal 6-Hydroxydopamine Parkinson's Disease Rat Model. Antioxidants (Basel). 12(6):1257.
- Li S, Rosen MC, Chang S, David S, Freedman DJ. (2023). Alterations of neural activity in the prefrontal cortex associated with deficits in working memory performance. Front Behav Neurosci. 17:1213435.
- Snytte J, Setton R, Mwilambwe-Tshilobo L, Natasha Rajah M, Sheldon S, et al. (2024). Structure-Function Interactions in the Hippocampus and Prefrontal Cortex Are Associated with Episodic Memory in Healthy Aging. eNeuro. 11(3):ENEURO.0418-23.2023.
- Yadav N, Toader A, Rajasethupathy P. (2024). Beyond hippocampus: Thalamic and prefrontal contributions to an evolving memory. Neuron. 112(7):1045-1059.
- Ogelman R, Gomez Wulschner LE, Hoelscher VM, Hwang IW, Chang VN, Oh WC. (2024). Serotonin modulates excitatory synapse maturation in the developing prefrontal cortex. Nat Commun. 15(1):1368.
- Karayol R, Medrihan L, Warner-Schmidt JL, Fait BW, Rao MN, Holzner EB, et al. (2021). Serotonin receptor 4 in the hippocampus modulates mood and anxiety. Mol Psychiatry. 26(6):2334-2349.